Вход / Регистрация
25.04.2024, 12:58
Выбор фона:



/ Новости сайта / Наука и Технологии / Предки китайцев и японцев скрещивались с двумя разными популяциями денисовцев
12.12.2018

Предки китайцев и японцев скрещивались с двумя разными популяциями денисовцев
Благодаря достижениям палеогеномики стало известно, что предки современных людей скрещивались с неандертальцами и денисовцами. Сегодня следы гибридизации с вымершими популяциями можно обнаруживать и без использования палеогенетических данных. Применив новый метод поиска чужеродных фрагментов ДНК к геномам 5639 современных жителей Евразии и Океании, американские генетики показали, что эпизодов гибридизации сапиенсов с денисовцами было не менее двух. При этом в геномах восточноазиатов есть следы обоих эпизодов, а у южноазиатов и папуасов — только одного. Две денисовские популяции, скрещивавшиеся с сапиенсами, различались по степени родства с алтайским денисовцем, чей геном пока является единственным прочтенным геномом денисовского человека.
Поздние представители рода Homo, такие как сапиенсы, неандертальцы и денисовцы, скитаясь по просторам Старого Света, неоднократно скрещивались друг с другом. Специалисты по палеогенетике и сравнительной геномике постепенно расшифровывают последовательность эпизодов гибридизации и направленность генетических потоков.
Поздние представители рода Homo, такие как сапиенсы, неандертальцы и денисовцы, скитаясь по просторам Старого Света, неоднократно скрещивались друг с другом. Специалисты по палеогенетике и сравнительной геномике постепенно расшифровывают последовательность эпизодов гибридизации и направленность генетических потоков.
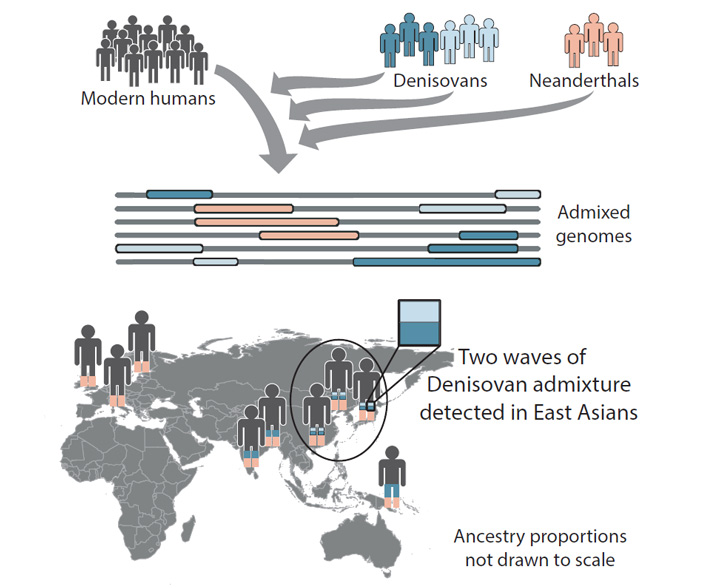
Рис. 1. «Графическое резюме» обсуждаемой статьи в Cell. Новый метод выявления чужеродных генетических примесей позволил установить, что предки современных жителей Восточной Азии скрещивались с представителями двух разных популяций денисовского человека. В геномах жителей Южной Азии и Океании есть следы скрещивания только с одной из них.
На сегодняшний день наиболее убедительно подтверждаются пять эпизодов гибридизации (рис. 2):
1) приток генов от неидентифицированных архаичных людей (возможно, поздних эректусов) к денисовцам,
2) приток генов от древних сапиенсов (вышедших из Африки ранее 60 000 лет назад) к алтайским неандертальцам,
3) от алтайских неандертальцев — к денисовцам,
4) от неандертальцев — к вышедшим из Африки сапиенсам около 55 000 лет назад (благодаря этому эпизоду в геномах внеафриканских сапиенсов присутствует около 2% неандертальской ДНК),
5) от денисовцев — к предкам современных жителей Австралии и Океании, а также, в меньшей степени, к предкам современного населения Южной и Восточной Азии.

Рис. 2. Эпизоды гибридизации, более или менее надежно установленные к 2016 году. Рисунок из статьи M. Kuhlwilm et al., 2016. Ancient gene flow from early modern humans into Eastern Neanderthals (см.: У алтайских неандертальцев найдены гены архаичных сапиенсов, а у денисовцев — гены гейдельбергских людей, «Элементы, 25.02.2016).
До сих пор не было ясности в вопросе о том, была ли гибридизация с денисовцами единичным событием.
Наверняка были и другие эпизоды. В частности, ряд фактов указывает на повторные скрещивания предков современных европейцев и азиатов с неандертальцами (Q. Fu et al., 2015. An early modern human from Romania with a recent Neanderthal ancestor; B. Vernot et al., 2016. Excavating Neandertal and Denisovan DNA from the genomes of Melanesian individuals). В геномах некоторых африканских народностей, таких как пигмеи, хадза и сандаве, обнаружены следы гибридизации с неизвестной архаичной популяцией, которая, по-видимому, отделилась от предков сапиенсов примерно тогда же, когда разделились линии сапиенсов и общих предков неандертальцев и денисовцев (J. Lachance et al., 2012. Evolutionary History and Adaptation from High-Coverage Whole-Genome Sequences of Diverse African Hunter-Gatherers; P. Hsieh et al., 2016. Model-based analyses of whole-genome data reveal a complex evolutionary history involving archaic introgression in Central African Pygmies).
Чужеродные примеси в геномах современных людей можно искать несколькими способами. Во-первых, можно искать фрагменты ДНК, совпадающие с известными неандертальскими и денисовскими последовательностями. Но тогда мы не сможем обнаружить примеси, источником которых были вымершие популяции, по которым нет палеогенетических данных. Чтобы найти такие примеси, используют статистические методы, не принимающие во внимание последовательности древних геномов. Эти методы основаны на том, что привнесенные (интрогрессированные, см. Introgression) фрагменты ДНК должны иметь определенное распределение длин (чем больше времени прошло с момента гибридизации, тем меньше их средняя длина) и содержать наборы тесно сцепленных (расположенных по соседству) редких полиморфизмов. Если речь идет о гибридизации, которая происходила уже после того, как предки разных рас и народов начали разбредаться по миру, то интрогрессированные участки должны встречаться только у некоторых, но не у всех современных популяций (например, неандертальские примеси есть у всех внеафриканских сапиенсов, но отсутствуют у африканцев).
Данный подход, известный под названием «S*-статистика», разрабатывается и совершенствуется уже более 10 лет (см.: V. Plagnol, J. D. Wall, 2006. Possible Ancestral Structure in Human Populations; B. Vernot, J. M. Akey, 2014. Resurrecting surviving Neandertal lineages from modern human genomes).
Американские генетики разработали новую, улучшенную версию S*-статистики, которую они назвали Sprime (по этой ссылке доступно программное обеспечение). Метод Sprime позволяет анализировать сразу большие участки генома (вместо последовательного обсчета коротких участков) и учитывать данные сразу по множеству геномов (вместо попарных сравнений). Кроме того, Sprime, в отличие от прежних версий, успешно справляется с таким осложняющим обстоятельством, как ограниченная миграция носителей интрогрессированных фрагментов ДНК в районы, населенные людьми без таких фрагментов. Например, Sprime позволил бы найти неандертальские примеси у неафриканцев, даже если бы некоторое (не слишком большое) число носителей этих примесей в далеком прошлом вернулось в Африку и скрестилось с местным населением. Конечно, без привлечения палеогенетических данных Sprime не определил бы, что это именно неандертальские примеси, но он бы показал, что это привнесенные фрагменты ДНК.
Проверив новый метод на виртуальных (сгенерированных компьютером) геномах с известной историей, исследователи убедились, что он обладает повышенной надежностью и чувствительностью по сравнению с прежними версиями. Так, если две популяции разошлись сотни тысяч лет назад, период гибридизации был десятки тысяч лет назад, и при этом в генофонд изучаемой популяции попало около 3% чужих генов, то метод Sprime позволит обнаружить примерно половину интрогрессированных фрагментов (остальные окажутся слишком короткими и/или содержащими слишком мало полиморфизмов, уникальных для второй популяции), а доля ложноположительных результатов будет не более 7%.
Убедившись, что метод работает, авторы применили его к 5639 полным геномам современных людей со всех концов Евразии. В выборку были включены также папуасы и африканцы йоруба (последние использовались в качестве контроля, поскольку считается, что у них практически нет неандертальских и денисовских примесей).
Количество и суммарная длина идентифицированных чужеродных фрагментов в геномах людей из разных популяций совпали с полученными ранее оценками доли неандертальской и денисовской ДНК в этих популяциях (с учетом того, что метод Sprime, как говорилось выше, обнаруживает примерно половину интрогрессированных участков). В частности, подтвердилось, что у восточноазиатов примесей больше, чем у европейцев и южноазиатов.
Напомним, что все эти чужеродные фрагменты были выявлены при помощи метода Sprime без использования палеогенетических данных. Теперь, конечно, нужно было их сравнить с известными древними геномами. Авторы так и поступили, использовав для сравнения два наиболее качественно прочтенных генома — алтайского неандертальца и денисовца. Анализировались только те интрогрессированные участки, в пределах которых и у неандертальца, и у денисовца есть как минимум по 10 однонуклеотидных отличий от современных людей (точнее, от африканцев, у которых нет неандертальских и денисовских примесей). Это дало возможность для каждого интрогрессированного участка определить степень его сходства, с одной стороны, с алтайским неандертальцем, а с другой — с денисовцем. При оценке сходства учитывались только те позиции, по которым неандертальцы и (или) денисовцы отличаются от африканцев.
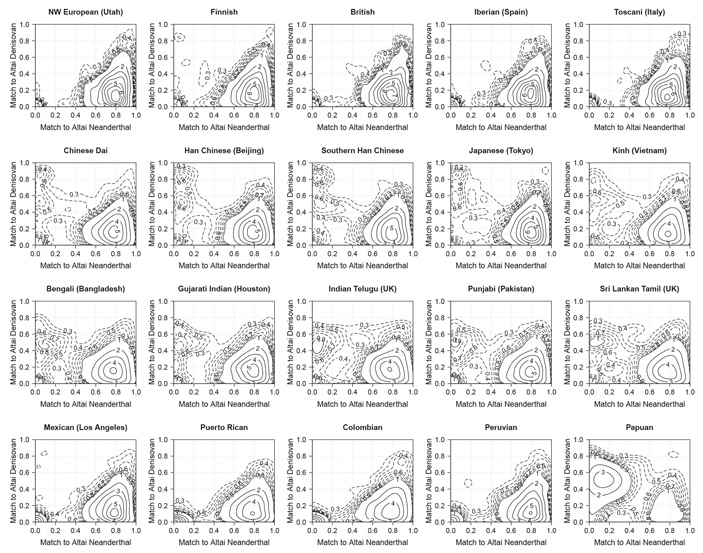
Рис. 3. Диаграммы, показывающие количество интрогрессированных фрагментов ДНК с разным уровнем сходства с геномом алтайского неандертальца (по горизонтальной оси) и денисовца (по вертикальной оси). Популяции, представленные на диаграммах 16–19 (от мексиканцев до перуанцев) — это не индейцы, а люди смешанного, преимущественно европейского происхождения. Прочие пояснения в тексте. Рисунок из обсуждаемой статьи в Cell
Полученные результаты представлены на рис. 3. На диаграммах, построенных для 20 внеафриканских популяций, показана частота встречаемости интрогрессированных фрагментов ДНК с разным уровнем сходства с геномами алтайского неандертальца и денисовца. Эти диаграммы заслуживают внимательного разглядывания.
На всех диаграммах мы видим маленькую компактную область с высокими значениями в левом нижнем углу. Это фрагменты, которые были определены программой Sprime как интрогрессированные, но которые при этом не имеют ничего общего ни с неандертальской, ни с денисовской ДНК. Скорее всего, это ложноположительный сигнал, то есть фрагменты, которые на самом деле не являются интрогрессированными и были определены программой ошибочно. Судя по результатам моделирования, программа должна давать 5–7% ложноположительных результатов, что и подтверждается.
На всех диаграммах мы также видим внушительный подъем в правом нижнем углу. Это — наше неандертальское наследие. Последовательности, унаследованные от неандертальцев, сходны с геномом алтайского неандертальца в среднем на 80%. При этом сходство с денисовцем у них низкое — порядка 10–20%. Здесь нужно пояснить, что денисовцы чуть ближе к неандертальцам, чем к сапиенсам, а это значит, что общие предки денисовцев и неандертальцев успели накопить какое-то количество общих отличий от сапиенсов еще до того, как разделились. Именно поэтому сходство неандертальских фрагментов с денисовским геномом достоверно отличается от нуля.
Денисовских примесей среди исследованных популяций больше всего у папуасов (последняя диаграмма). Средний уровень сходства денисовских включений в папуасских геномах с известным геномом денисовца — около 50% (а с геномом алтайского неандертальца — те же 10–20%). Это значит, что те денисовцы, которые оставили свой след в геномах папуасов, были довольно дальней родней девочки из Денисовой пещеры, чей геном удалось отсеквенировать. Они представляли другую популяцию денисовцев, которая обитала, возможно, далеко от Алтая (может быть, где-то на юге или юго-востоке Азии).
В геномах жителей южной Азии (третий ряд диаграмм: индусы, бенгальцы и др.) тоже есть денисовская примесь, хоть и не такая большая. При этом уровень сходства привнесенных участков с геномом денисовской девочки такой же, как у папуасов: около 50%. Это значит, что источником примеси, вероятно, была та же самая популяция денисовцев, не слишком близкая к алтайской, что и у папуасов.
В геномах европейцев (первый ряд диаграмм) денисовских примесей практически нет, за исключением финнов (диаграмма №2), у которых есть чуть-чуть денисовской ДНК. Скорее всего, эта примесь попала к финнам вместе с порцией азиатских генов (M. Sikora et al., 2014. Population Genomic Analysis of Ancient and Modern Genomes Yields New Insights into the Genetic Ancestry of the Tyrolean Iceman and the Genetic Structure of Europe).
Самый интересный и неожиданный результат связан с восточноазиатами (три китайских выборки и одна японская, диаграммы 6–9 во втором ряду). Их денисовская примесь состоит из двух разнородных частей: одни денисовские фрагменты сходны с геномом алтайской девочки на 50% (как у папуасов и южноазиатов), а другие — примерно на 80%. Двугорбость распределения статистически достоверна. Это значит, что у предков китайцев и японцев, скорее всего, было два эпизода гибридизации с денисовцами. Один раз они скрещивались с той же популяцией, что оставила след в геномах папуасов и южноазиатов, а второй — с другими денисовцами, более близкими к алтайской популяции.
Эпизод с участием «50-процентных» денисовцев мог быть тем же самым, что и у предков папуасов и южноазиатов. Иными словами, в этом эпизоде могли участвовать еще общие предки папуасов и жителей южной и восточной Азии. Или, может быть, собственно с денисовцами скрещивались только предки папуасов, а уже потом от них, в результате каких-то миграций, получили денисовскую примесь предки индусов и китайцев. Что же касается эпизода с участием «80-процентных» денисовцев, то в нем были задействованы только предки восточноазиатов.
Определить временную последовательность эпизодов гибридизации в принципе можно по длине интрогрессированных фрагментов. В геномах восточноазиатов неандертальские фрагменты в среднем чуть короче, чем «50-процентные» денисовские, а те, в свою очередь, короче, чем «80-процентные». Это указывает на возможную последовательность событий: сначала — скрещивание с неандертальцами, потом — с «50-процентными» и, наконец, с «80-процентными» денисовцами. Однако авторы по-честному отмечают, что различия по длине фрагментов не являются статистически достоверными, и поэтому делать какие-то выводы на их основе преждевременно.
Новые данные не подтвердили точку зрения о множественных эпизодах гибридизации с неандертальцами. Неандертальские фрагменты ДНК не распадаются ни на какие кластеры, а это значит, что либо эпизод был только один, либо повторные скрещивания происходили с неандертальскими популяциями, очень близкими к той, с которой предки внеафриканских сапиенсов гибридизовались изначально около 55 000 лет назад. Второй вариант более вероятен, потому что у азиатов достоверно больше неандертальской ДНК, чем у европейцев, и это не удается объяснить действием отбора (то есть тем, что у предков европейцев отбор эффективнее вычищал неандертальскую примесь). Скорее всего, предки азиатов повторно скрещивались неандертальцами, которые были близкой родней тех, с которыми ранее скрещивались общие предки всех внеафриканских сапиенсов.
Авторы также посмотрели, какие из идентифицированных чужеродных фрагментов ДНК подвергались действию положительного отбора, то есть пошли на пользу нашим предкам и потому достигли высокой частоты в современных генофондах. Подтвердились прежние выводы, согласно которым отбор поддержал целый ряд неандертальских аллелей, связанных со строением кожи, волос и, главное, с иммунной системой. Это логично, потому что вышедшие из Африки сапиенсы столкнулись в Евразии с новыми патогенами, так что гены иммунной защиты местного населения могли оказаться очень кстати. От денисовцев наши предки получили меньше полезных генетических вариантов, чем от неандертальцев (см.: Наши предки заимствовали у неандертальцев и денисовцев важные гены для защиты от вирусов, «Элементы», 07.10.2011; Тибетцы унаследовали от денисовских людей ген, спасающий от гипоксии, «Элементы», 10.07.2014).
Таким образом, благодаря развитию методов палеогенетики и биоинформатики (анализа геномных последовательностей) наши знания об истории гибридизации сапиенсов с «архаичными Homo» быстро уточняются и детализируются. Прогресс не может не впечатлять, особенно если вспомнить, что с того момента, когда сам факт такой гибридизации был впервые твердо установлен, прошло менее восьми лет.
Источник: http://elementy.ru
Комментарии 4

|
0 |

|
0 |



|
0
Partizan-16
12.12.2018 18:05
[Материал]
Люди ни как не могли сами скрещиваться с другими видами. Гориллы или мартышки с орангутанами думаю мало кого привлекают. Наги намеренно создавали разные популяции людей из всего, что под руку попало.
|
Разговоры у камина

Новое на форуме
Последние комментарии
Архив записей
Статистика
Мы в соцсетях

